S961 Mediates the Inhibition of Hyperinsulinemia-Induced Growth in Androgen-Independent PC-3 Cells
Keywords: Insulin resistance, Hyperinsulinemia, Prostate cancer, PC-3 cells, High-fat diet, S961
Abstract
Through the application of the S961 insulin receptor antagonist, we investigated the effect of insulin and diet-induced hyperinsulinemia on the growth of the PC-3 cells in vitro and discerned the growth promoting effect of insulin in the androgen-independent cells. Sprague-Dawley rats were kept on a high-fat diet for 4 weeks for the induction of insulin resistance and hyperinsulinemia. Insulin alone or serum of the rats kept on either normal-pellet diet or high-fat diet was used to stimulate the serum-starved PC-3 cells growth in culture. The selective S961 peptide, a high-affinity insulin-receptor antagonist, was used to confirm the insulin-mediated effects. Significant impairment in the glucose disposal rate and increase in the serum glucose and insulin levels was observed in the high-fat-diet-fed rats. The media supplemented with the serum of the high-fat-diet-fed rats accelerated the growth of the PC-3 cells in comparison to that of normal-pellet-diet-fed rats. Insulin treatment led to accelerated growth of the serum-starved PC-3 cells in a dose-dependent manner and was significantly inhibited by the S961 pretreatment. Insulin and serum of the diet-induced hyperinsulinemic rats promote the growth of androgen-independent prostate cancer PC-3 cells. Further, our results provide support for the concept that diet-associated elevation in insulin level may augment the growth of prostate cancer cells.
Introduction
Insulin resistance is a highly prevalent condition all over the world, characterized by impairment in insulin-mediated effects and compensatory rise in insulin level. Epidemiological studies suggest that insulin resistance increases the risk of numerous cancers. A recent prospective cohort study involving more than 90,000 men and women revealed an association between obesity and mortality from different malignancies. To explain this association, several plausible mechanisms have been proposed, such as the effect of metabolic syndrome on sex hormones, hyperinsulinemia, increased tumor growth, and the chances of underdosing with chemotherapeutic agents.However, the hyperinsulinemic state, an important consequence of insulin resistance, appears to be the most appropriate explanation for the association between metabolic syndrome and cancer. High levels of circulating insulin can promote tumor growth by overactivating the Insulin-like growth factor 1 receptor pathway, which drives cellular survival and inhibits apoptosis.
Interventions such as low-fat diet and/or strenuous exercise are reported to reduce growth and induce apoptosis in prostate cancer cells. Furthermore, in a recent study, Vankateswaran et al. reported accelerated growth of androgen-dependent LnCap tumors in the diet-induced hyperinsulinemic mice. These experiments provide convincing evidence that a low-fat, high-fiber diet and daily exercise, which are associated with decreases in insulin level, reduce the growth of prostate cancer. However, the effect of insulin and insulin-resistance-associated hyperinsulinemia on the growth of androgen-independent prostate cancer is less clear.
The present study aimed to investigate the role of insulin and diet-induced hyperinsulinemia on the growth of androgen-independent prostate cancer. Insulin/insulin-like growth factor-1 (IGF-1) signaling is known to activate androgen signaling through direct interaction of Foxo-1 with androgen receptor. Therefore, the PC-3 cells that do not express androgen-receptor and are unresponsive to androgen treatment have been selected to examine the direct growth-promoting effect of insulin. We have examined the growth of PC-3 cells in response to insulin alone or serum of diet-induced hyperinsulinemic rats. The results of the present study clearly indicate that insulin and diet-induced hyperinsulinemia accelerate the growth of androgen-independent PC-3 cells.
Materials and Methods
Chemicals
D-glucose and insulin were procured from Sigma-Aldrich (St. Louis, MO). Ham-12 media, fetal bovine serum (FBS), and L-glutamine were procured from Hi-media (Mumbai, India). Insulin receptor antagonist S961 was procured from Novo-Nordisk (Bagsvaerd, Denmark). Lyophilized S961 was dissolved in sterilized phosphate buffered saline (PBS) and stored in aliquots at negative 20 degrees Celsius, freshly prepared by diluting appropriately to achieve desired concentration with Ham-12 media just before use.
Cell Culture
PC-3 cells were obtained from National Centre for Cell Sciences (Pune, Maharastra, India) and were maintained in Ham-12 media supplemented with 10% fetal bovine serum (FBS) at 37 degrees Celsius in a humidified incubator chamber containing 5% carbon dioxide. All the experiments were performed between passage number 30 to 33. Effect of different treatments on the growth of PC-3 cells was assessed by using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay in 2 to 3 replicates.
Approximately 7,500 cells were seeded in the 96-well, flat-bottomed plates in 10% FBS containing Ham 12 media and incubated overnight at 37 degrees Celsius. The FBS containing media was removed and cells were kept on 24-hour serum starvation in Ham 12 media. The cells were exposed to the different treatments in a serum-free Ham 12 media and incubated for 24 hours. The medium was removed, replaced with 100 microliters of MTT (0.5 mg/ml) in 10% FBS containing Ham 12 media, and cells were incubated at 37 degrees Celsius for 4 hours. Supernatant was removed from the wells and the reduced MTT dye was dissolved in 200 microliters per well DMSO. Absorbance was determined at 550 nm.
Further, the effect of insulin on the growth of PC-3 cells was ascertained by direct counting of the cells. Briefly, 20,000 cells were seeded in 24-well, flat-bottomed plates in 10% FBS containing Ham 12 media, and incubated overnight at 37 degrees Celsius. The FBS containing media was removed and cells were kept on 24-hour serum starvation in Ham 12 media. The cells were exposed to the different treatments in a serum-free Ham 12 media and incubated for 24 hours. The medium was removed and exposed to 200 microliters of 0.25% Trypsin-EDTA solution for 15 minutes, followed by 800 microliters of serum containing Ham-12 media. Cell suspension was loaded on each side of the Neubaurs chamber and cells were counted in 4 square millimeter area under an inverted microscope at 100 times magnification.
Acridine Orange and Ethidium Bromide Staining
After the treatment protocol, the cells were rinsed twice with cold PBS and resuspended at 1.5 times 10 to the 5 cells per milliliter in Ham 12 media containing 15% FBS. Ten microliters of dye mixture (100 micrograms per milliliter Acridine orange and 100 micrograms per milliliter Ethidium bromide) were added to the 100 microliters of cell suspension. After gentle agitation, 20 microliters of cell suspension-dye mixture was placed on the slide and covered with a coverslip. A minimum of 400 cells were examined under a fluorescent microscope. The cells were scored as follows: normal cells with bright green chromatin, apoptotic cells with orange chromatin, and necrotic cells with red chromatin.
Determination of the Effect of Diet-Induced Insulin Resistance on the Growth of PC-3 Cells
All the animal experiments were approved by the Institutional Animal Ethics Committee and were used according to the Committee for the Purpose of Control and Supervision of Experiments on Animals (CPCSEA) guidelines. Male Sprague-Dawley rats (200 to 220 grams) were procured from Institute’s Central Animal Facility and kept at controlled environmental conditions and were allowed to access the food and water ad libitum.
Insulin resistance was induced in rats by feeding a high-fat diet (HFD), whereas the control rats were fed with a normal-pellet diet (NPD) for a period of 4 weeks. The NPD used to feed the animals was standard rodent chow (Pranaw Agro Industries, New Delhi, India). The detailed methodology for the HFD preparation has already been described by Srinivasan et al. In brief, the content of HFD diet includes NPD powder (36.5%), lard (31%), casein (25%), vitamin-mineral mix powder (6%), cholesterol (1%), DL-methionine (0.3%), Yee-sac powder (0.1%), and sodium chloride (0.1%).
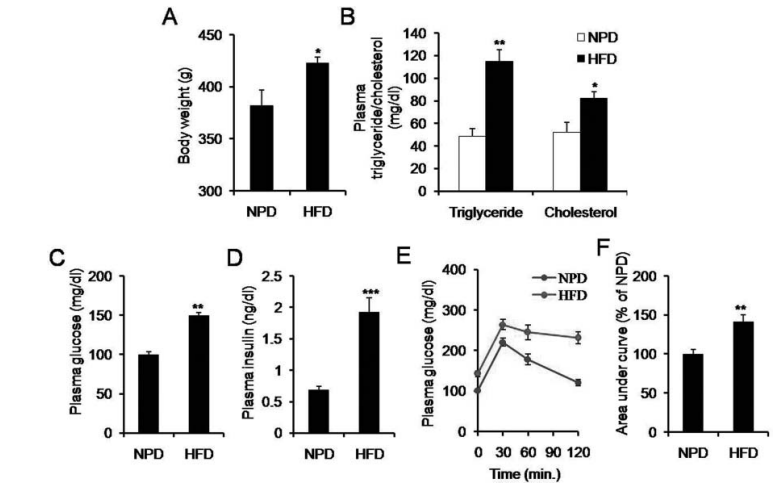
Figure 1 shows that a high-fat diet (HFD) induces insulin resistance and hyperinsulinemia. Panels A and B show that HFD-fed rats had significantly higher body weight, plasma triglyceride, and cholesterol levels compared to rats fed a normal-pellet diet (NPD). Panels C and D show increased plasma glucose and insulin levels in HFD-fed rats. Panel E shows impaired glucose disposal in HFD-fed rats based on an intraperitoneal glucose tolerance test. Panel F shows a significant increase in the area under the curve for this test in HFD-fed rats. Data are presented as mean ± SEM. **P < 0.01, ***P < 0.001 vs. NPD.
Induction of insulin resistance and hyperinsulinemia was confirmed by intraperitoneal glucose tolerance test (IPGTT), examining the hepatic histology and measuring the level of glucose and insulin in the serum as described previously. The fasting (6 hours) blood of NPD-fed and HFD-fed rats was collected from the retro-orbital plexus under light ether anesthesia, allowed to clot for 30 minutes, and serum was collected after centrifugation (5,000 rpm times 5 minutes). Serum was filtered using a sterile syringe filter (0.22 millimeter) for in vitro experiments.
Approximately 7,500 cells were seeded in 96-well, flat-bottomed plates in 10% FBS containing Ham 12 media and incubated overnight at 37 degrees Celsius. The FBS containing media was removed, and after 24-hour serum starvation, cells were exposed to the Ham 12 media with or without 10% FBS, 10% serum of NPD-fed rats, or 10% serum of HFD-fed rats and incubated for 1, 3, or 5 days. The growth of cells was determined by MTT assay as described previously. The reading taken from 1 control plate just after the serum starvation on day 2 was considered as the base value to calculate the cell growth in response to different growth conditions.
Statistical Analysis
Statistical analysis was performed using Jandel SigmaStat statistical software. Significance of difference between 2 groups was evaluated using Student’s t-test. For multiple comparisons, ANOVA was used and post hoc analysis was performed with Tukey’s test. Results were considered significant if P values were less than or equal to 0.05.
Results
Effect of HFD Feeding
HFD feeding led to a significant increase in body weight, plasma triglyceride, and plasma cholesterol level. To examine the effect of HFD on insulin sensitivity, plasma insulin level and impairment in the glucose tolerance was examined. Significant (P less than 0.01) increase in plasma insulin (1.9 plus or minus 0.2 vs. 0.7 plus or minus 0.1 ng per dl) and glucose (149.3 plus or minus 4.5 vs. 100.0 plus or minus 3.4 mg per dl) level was observed in the HFD-fed rats as compared to the NPD-fed rats. Furthermore, HFD-feeding led to hepatic deposition of fat and impairment in the glucose disposal as compared to NPD-fed rats.
Effect of Diet-Induced Insulin Resistance on the Growth of PC-3 Cells
The media supplemented with the FBS, serum of NPD-fed rats, or serum of HFD-fed rats accelerated the growth of PC-3 cells. However, the media supplemented with the serum of HFD-fed rats induced significant increase in the growth of the PC-3 cells in comparison to the cells grown in the media supplemented with the serum of NPD-fed rats.
Effect of Insulin on the Growth of PC-3 Cells
Insulin treatment led to significant increase in the growth of 24-hour serum-starved PC-3 cells in a dose-dependent manner as determined by manual counting of cell population and MTT assay. However, no appreciable change in the apoptosis index was observed in response to the insulin treatment. Pretreatment with insulin-receptor antagonist S961 significantly inhibited insulin-induced growth of the PC-3 cells.
Discussion
Prostate cancer, the second most common cancer in men, and insulin resistance are major health concerns worldwide. Previous studies have demonstrated an association between metabolic syndrome and various cancers, with a particular emphasis on colon, endometrial, liver, and pancreatic cancers. Although experimental and clinical studies support the connection between dietary habits, metabolic syndrome, and prostate cancer, the precise understanding of the underlying mechanisms are not completely known. Furthermore, recent literature proposes insulin resistance associated secondary hyperinsulinemia as an important factor for the progression of prostate cancer. Results of the present study clearly demonstrate that insulin and diet-induced hyperinsulinemia accelerates the growth of the human androgen-independent PC-3 cells.
In addition to playing a critical role in the glucose homeostasis, insulin has a growth-stimulating effect. Insulin receptors are ubiquitously expressed in normal as well as malignant tissues. The MEK ERK pathway of insulin signaling is primarily concerned with the growth-stimulating effects of the hormone. PC-3 is a poorly differentiated prostate cancer cell, derived from the bone metastasis of human prostatic adenocarcinoma, unresponsive to androgen treatment in vitro and does not express androgen receptor. Recently, Fan et al. reported that insulin/IGF-1 signaling activates androgen signaling through direct interaction of Foxo 1 with the androgen receptor. PC-3 cells, which do not express androgen receptor, provide an ideal in vitro test system to examine the direct growth-promoting effects of insulin.
In the present investigation, insulin accelerated the growth of serum-starved PC-3 cells in a dose-dependent manner. Further, the growth promoting effect of insulin is confirmed by insulin receptor antagonist S961. Pretreatment with S961 inhibited insulin-induced growth of PC-3 cells. S961 is a highly selective peptide antagonist to the insulin receptor and reported to block insulin mediated effects in vitro as well as in vivo.
Insulin resistance is an increasingly prevalent condition all over the world and is often complemented with a compensatory rise in the plasma insulin level. Hammarsten et al. reported that hyperinsulinemia promotes prostatic growth and is an important risk factor for prostate cancer. Previously, it has been reported that a low-fat diet or intensive exercise results in the induction of apoptosis and reduced plasma insulin, IGF-1 level, and growth of prostate cancer (LnCap) and primary prostate epithelial cells in vitro. Further, diet-associated hyperinsulinemia has been reported to accelerate the growth of LnCap cells in vivo. Elevated serum levels of insulin within normal range has been found to be associated with a higher risk of prostate cancer. In a recent study, Hammersten et al. reported age, type 2 diabetes, dyslipidemia, hyperinsulinemia, and metabolic syndrome as prospective risk factors for lethal clinical prostate cancer.
To examine the effect of insulin resistance on the growth of PC-3 cells, the cells were grown in Ham-12 media supplemented with the serum of either insulin-resistant or insulin-sensitive rats. Ham-12 media supplemented with the serum of insulin-resistant rats induced significant increase in the growth of the PC-3 cells as compared to that of NPD-fed rats. The level of insulin in the serum of HFD-fed rats was approximately threefold higher than that of NPD. Thus, the increased cell growth in the presence of the serum of HFD-fed rats can be attributed to the increased insulin level.
However, there is a possibility that apart from elevation in the insulin level and other factors such as change in the IGF-1 axis, hyperlipidemia may also contribute to the growth of the PC-3 cells. The crucial role of IGF-1 signaling is well established in the progression of prostate cancer, and it is considered a link between dietary habit and the disease. Furthermore, dietary fat is known to alter IGF-1 and/or IGFBP-1 levels and thus have a direct as well as indirect effect on the free IGF-1 level. Ngo et al. reported that individuals kept on a low-fat diet and exercise intervention exhibited approximately 20% decrease in IGF-1 and 53% increase in IGFBP-1 in the serum. Further, the serum of humans kept on low-fat diet and exercise intervention exerted less growth promoting effects (approximately 30%) on the androgen-dependent LnCaP cells.
Mavropoulos et al. examined the effect of varying carbohydrate and fat content on survival in a murine LnCaP prostate cancer xenograft model and concluded that dietary carbohydrate restriction favorably alters the IGF axis and might slow the progression of the prostate cancer. However, twice-a-week intermittent fasting in mice kept on a Western diet did not produce any beneficial effects. Although, IGF-1 is having a relatively stronger mitogenic effect in comparison to insulin, the chronic increase in insulin might have important implications in the regulation of cellular proliferation.
Moreover, the hyperlipidemic condition might affect the prostatic cell proliferation through activation of the inflammatory autotoxin-lysophosphatidic acid (ATX-LPA) pathway and/or PPAR alpha/gamma signaling. Increased oxidative stress and activation of the NF-kB in the prostate of the HFD-fed mice indicates the crucial role of inflammation in the prostatic cell proliferation. Further, chronic HFD-feeding is associated with low-grade systemic inflammation and raises the possibility of the presence of inflammatory mediators in the serum and subsequent influence on the growth of the PC-3 cells.
A series of studies conducted by W. J. Aronson and coworkers have provided valuable information about the effect of lifestyle modifications such as change in dietary constituents and the intervention of exercise on the growth characteristics of prostate cancer cells. Summing together the literature reports and the results of the present study, it clearly appears that the HFD can exert its growth-promoting effects through multiple pathways, such as activation of insulin signaling, IGF-1 signaling, PPAR alpha/gamma signaling, and ATX-LPA axis.
The results of the present study clearly indicate that insulin accelerates the growth of the serum-starved androgen independent human prostate cancer (PC-3) cells. Furthermore, the serum of diet-induced insulin-resistant rats promotes the growth of the PC-3 cells and supports the hypothesis that insulin resistance associated compensatory hyperinsulinemia is an important risk factor for prostate cancer. However, further studies investigating the presence of different inflammatory mediators, the interaction between inflammatory pathways and insulin signaling, and pathway-specific (IRS PI-3Kinase or MEK ERK) inhibition of insulin-receptor signaling are needed to better understand the involvement of insulin in the development and progression of prostate cancer.